 Four-wing saltbush (Atriplex canescens), native to western North America, extends from Canada to Mexico and from the Great Plains to the Pacific Coast.
Four-wing saltbush (Atriplex canescens), native to western North America, extends from Canada to Mexico and from the Great Plains to the Pacific Coast.
Shrubby species of Atriplex are in the family Chenopodiaceae, which contains other important shrubs such as winter fat (Krascheninnikovia lanata), and often dominate landscapes in many arid and semi-arid regions, particularly in habitats that combine high soil salinity with aridity. Four-wing saltbush is an important browse species to wildlife and livestock and has been reported to provide as much as 11.4 % to 13.6% crude protein.
The use of four-wing saltbush in restoration and land rehabilitation plantings is well documented and increasingly popular. Four-wing saltbush is a common species to be seeded on Great Basin rangelands, especially in the more xeric Wyoming big sagebrush (Artemisia tridentata) and salt desert shrub plant communities.
Granivorous rodents are important in the ecology of plant communities as well as the management practices that occur in those communities. Granivorous rodents have been reported to be an important seed dispersal mechanism for numerous Great Basin plant species, such as antelope bitterbrush (Purshia tridentata) and Indian ricegrass (Achnatherum hymenoides), as well as an active seed and seedling predator on Great Basin plants species.
Granivorous rodents exhibit two types of seed caching behavior; they cache some seeds in larders deep within their burrows, referred to as “larder hoarding”, and cache some seeds in shallow depressions they dig throughout their home range, referred to as “scatter hoarding”.
Larder hoard caches are buried at depths that may allow germination but are most often too deep to sprout, whereas scatter hoard caches that are not recovered are buried at depths that often promote germination and therefore have been found to be an important mechanism for the recruitment of various range plants.
In any land management practice that involves seeding in restoration or rehabilitation efforts, land managers must be coherent about the role that biotic and abiotic factors ultimately have on the success and failures of these efforts. Four-wing saltbush has been reported to experience quite variable success in seeding efforts as poor seed germination or lack of proper amount and periodicity of precipitation would fit under the abiotic category and are more acknowledged. Biotic factors such as seed and seedling predation by granivorous rodents are less understood. Limited research has reported that the rodents seldom cache four-wing saltbush seed, while it has also been reported that rodents damage young four-wing saltbush plants.
To investigate the behavior and role granivorous rodents exhibit with four-wing saltbush we initiated a study to address: 1) The harvest, consumption, and caching of four-wing saltbush seed, and 2) the possible consumption of four-wing saltbush seedlings by granivorous rodents. We hypothesized that granivorous rodents in this study would harvest, consume and cache a portion of the four-wing saltbush seed they interacted with. We also hypothesized that four-wing saltbush seedlings would also be consumed by granivorous rodents.
First, we selected a study site east of Reno, NV, Desert Queen Valley, which is a salt desert shrub habitat dominated by four-wing saltbush, shadscale (Atriplex confertifolia), Bailey’s greesewood (Sarcobatus baileyi), winter fat, Indian ricegrass and Russian thistle (Salsola tragus). The site is very arid and only receives an average of 4” – 6” of annual precipitation.
To investigate rodent interaction with four-wing saltbush, we constructed portable live-trap enclosures where consumption and caching of four-wing saltbush seed and seedlings by various rodents could be recorded. These portable live-traps, 24” x 12” x 14”, were constructed having solid plywood bottoms, fronts and backs and were covered with 0.25” mesh hardware cloth (Figure 1).

A hole was cut out of the front plywood panel to accommodate the placement of a Sherman live-trap, with the back door of the live-trap removed, to allow rodents to enter but not exit the enclosure. The back plywood was removable to allow for the placement and removal of a 20” x 10” x 2” deep soil flat.
A total of 12 portable enclosures, 2 enclosures permitted no access (controls), were randomly placed out on a 12 transect by 10 station grid. Each transect and station was at 50’ spacing.
Enclosures were set out each evening at 1600h and baited with 15 ounces of millet seed (600 – 650 seeds) in the live-trap. One hundred four-wing saltbush seeds, collected from the site the previous year, were placed on the surface of the soil in the soil flats. Soil in the soil flat was collected at the site and sieved free of any seed. Enclosures were checked the following morning at 600h.
Enclosures with a rodent capture were dealt with first. Rodents were recorded by identification, sex, and tagged with a numbered ear tag, or in the case of a recapture the ear tag number was recorded. Cheek pouches were checked for any seeds and removed and recorded. Rodents were released at the point of capture. The number of visible seeds on the surface of the soil was noted, then the soil flat was sieved, and the number of seeds recorded as un-cached, cached, or consumed. The portable enclosures were then reset and randomly placed out within the grid.
This process took place for 5 consecutive mornings over 4 separate weeks from late September through early November, the timing of four-wing saltbush seed fall in the area.
These portable enclosures were also used to investigate any four-wing saltbush seedling consumption by the various rodents at the site. Four-wing saltbush seedlings were grown in soil flats in soil from the site in a greenhouse environment. Twenty-five seedlings were grown in each flat and placed into the portable enclosures (Figure 2).

Using the same transects as in the seed part of the study, portable enclosures were placed out at 1600h each evening, baited with 15 ounces of millet seed inside the live-trap and checked the following morning at 600h. Consumption of four-wing saltbush seedlings was recorded as well as any damage to seedlings not consumed (e.g. digging up). New soil flats with fresh four-wing saltbush seedlings were placed inside the enclosures and reset at randomly selected stations within the grid. This process took place for 5 consecutive mornings for 4 separate weeks from late March through late May, the timing of four-wing saltbush seedling sprouting at the site.
A total of 5,400 trap nights were conducted during this study, resulting in a total of 132 captures of 59 separate individuals of 4 separate species. The species richness and diversity of the rodent population at this site, as represented through the live-trap data, yielded that the site was dominated by the Merriam’s kangaroo rat (Dipodomys merriami) (34), followed by the chisel-toothed kangaroo rat (Dipodomys microps) (13), Desert kangaroo rat (Dipodomys deserti) (8), and the White-tailed antelope ground squirrel (Ammospermophilus leucurus) (4). The trapping grid was 2.5 acres in size and therefore represents a minimum rodent population of 24/acre.
Investigation of rodents, 103, with four-wing saltbush seed in the portable enclosures revealed that none of the 103 rodents harvested four-wing saltbush seed in their cheek pouches or cached any seed in the portable enclosure. Therefore, there was no evidence that rodents were attempting to cache four-wing saltbush seed. However, all 103 rodents that entered the portable live-trap enclosures in this study did however harvest and store a portion of the millet seed available to them in their cheek pouches or mouth cavity 100% of the time.
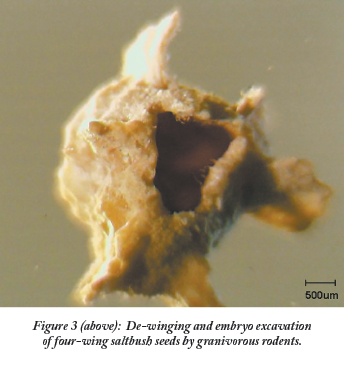
Although rodents did not harvest four-wing saltbush seed in their cheek pouches they did consume four-wing saltbush seed as they excavated the embryos from the seed coat (Figure 3).
The Merriam’s and Desert kangaroo rat as well as the White-tailed antelope ground squirrel significantly consumed four-wing saltbush seed. The Desert kangaroo rat consumed the most four-wing saltbush seed, 55.11 (± 3.22), n = 29, followed by the Merriam’s kangaroo rat, 54.70 (± 2.20), n = 62 (Figure 4), White-tailed antelope ground squirrel, 34.33 (± 8.14), n = 5, and the chisel-toothed kangaroo rat, 13.63 (± 6.60), n = 7.
There were no observations of the white-tailed antelope ground squirrel in our portable live-trap enclosures during the seedling interaction period of the study. The remaining 3 rodent species that entered the portable enclosures, 86, at the site significantly consumed four-wing saltbush seedlings (Figure 4).

The consumption of four-wing saltbush seedlings by these rodents was very high as the Desert kangaroo rat consumed an average of 99.3, n = 12, Merriam’s kangaroo rat, 92.9, n = 51, and the chisel-toothed kangaroo rat, 74.6, n = 23. Even though millet seed was available as an alternative food source, the 15 ounces of millet seed was available and never totally consumed.
As one may expect, the disturbance caused by individual rodents varies as some of the four-wing saltbush seedlings are trampled, therefore we only counted those seedlings that were physically apparent as being foraged upon and consumed.
From field observations, we suspected that four-wing saltbush seed would not be a preferred seed species for granivorous rodents as there is visual evidence of four-wing saltbush seed falling to the ground beneath the shrub and accumulating over-time, which is not the case with seeds from preferred shrub species such as antelope bitterbrush.
Four-wing saltbush seed has a wind dispersal mechanism as the seed physically contains large-winged appendages, whereas shrubs such as antelope bitterbrush seed does not have an active wind dispersal system and relies heavily on rodent dispersed seed. Although we hypothesize that rodent species tested in this study would harvest and cache a portion of the four-wing saltbush seed they interacted with, we did not record any harvest and caching behavior.
Even though the large wing structures of four-wing saltbush seed and the very hard seed coat would be considered deterrents for granivorous rodents, we observed a high degree of de-winged and embryo excavation on four-wing saltbush seed (Figure 3).
We also hypothesized that granivorous rodents would graze on four-wing saltbush seedlings which we did record high levels of this behavior. Our field experience has resulted in the observation of numerous species being grazed at the early seedling stages such as antelope bitterbrush, Indian ricegrass, and winter fat.
It is important that critical browse species, such as four-wing saltbush, are vigorous and healthy as to experience good flowering and produce substantial seeds to the environment. The more seeds that are available to germinate and emerge, the less the total effect of rodent predation on the recruitment of seedlings to sustain the plant population.
Although this research was conducted using portable live-trap enclosures, granivorous rodents in this study avoided caching four-wing saltbush seed while still excavating the embryo from the seed. The seedling predation may well be higher than in natural conditions, yet the fact that such a high level of seedling predation occurred when an alternative millet seed food source was available suggest a preference for these seedlings at this young phenology stage. This research yielded that four-wing saltbush seed is not highly preferred as the rodents at this site are not harvesting and caching the seed for future consumption.
The high consumption of four-wing saltbush seedlings at this site is alarming and may be an explanation of poor success at given sites following the seeding of four-wing saltbush. The clipping of seedlings by granivorous rodents is more detrimental to shrub seedlings than grass seedlings due to the removal of the hypocotyl or epicotyl.
In the past three decades, numerous years have resulted in large wildfires that have burned millions of acres. For example, in 1999, more than 1.8 million acres burned in Nevada alone, which resulted in the largest rehabilitation/restoration undertaking in state history. This rehabilitation/restoration effort resulted in the purchase of nearly 4.8 million pounds of grass, forb and shrub seed to be put on the burned rangelands.
Four-wing saltbush seed was second only to Wyoming big sagebrush of desired shrub species in the 98,000 acres to be drill seeded and the more than 370,000 acres to be aerial seeded. The importance of four-wing saltbush for wildlife and domestic livestock as well as its’ role in improving functional plant communities only enhances the desire to successfully rehabilitate/restore degraded rangelands.
The results from this study would suggest that resource managers cannot depend on granivorous rodents to harvest, disperse and cache four-wing saltbush seed, therefore drill seeding of four-wing saltbush seed may be necessary to get the seed into the ground where the seed has a higher chance of germination and emergence. The fact that four-wing saltbush seedlings were consumed at a high level allows resource managers the understanding of that perhaps transplanting of older seedlings could result in less seedling predation.
Understanding the possible effects of rodent behavior with four-wing saltbush seed and seedlings should help resource managers in their planning and implementation of future rehabilitation/restoration efforts.
By Charlie D. Clements and Dan N. Harmon