On the Ground
- More often than not, there is untapped potential for win-wins between livestock production and conservation. On the other hand, it is impossible to achieve every objective everywhere, all the time. Sometimes the tradeoffs are real.
- We need to spend less time searching for general rules and more time embracing the complexity and context-dependence within rangeland science.
- Rather than writing off findings that do not fit our current worldview, we should challenge ourselves to broaden our views in ways that reconcile multiple findings or multiple truths. It is possible we are all partly or mostly right, and we just need to figure out why, how, and in what contexts.
- There is value in doing research in a way that focuses on really listening to and respecting multiple perspectives so that the results we produce not only qualify as facts, but also as truths that many people can buy into and get behind.
Introduction
Someone once told me that if you do not change your mind about something important every 10 years, you are stagnating as a human being. What you change your mind about cannot be something small. It has got to be something fundamental, something all your friends know about you. In other words, to keep growing as humans, we must transform.
In my own life and career, one of my goals has been to stay fiercely open-minded and be ready for transformation. I have always been tempted to resist categories, to zoom in at the boundaries and search for gradients and complexity. Here, I am going to tell you three stories about things I have changed my mind about in my relatively short (~15 year) career. I hope these stories will engage your curiosity or spark a discussion with your colleagues. I also hope these ideas will point to some strategies for moving through the controversies and challenges that face the rangeland community.
False Dichotomies
I am going to tell you something I really do not want you to know about me. But I put the word “humility” in my title, so this is where I have to start. I grew up in suburbia. I did not attend FFA—I did not even know what FFA was. My great uncle ran a small family dairy farm in upstate New York, so I thought all cows lived in big red barns with kittens in the loft. I knew wild animals lived in zoos and national parks, and other places where there were no people. I am telling you this because I want to emphasize that the people who work in or study rangelands come from a wide variety of backgrounds and perspectives. I am also telling you because, growing up in suburbia, I decided I was going to save all the world’s beautiful and diverse plants and animals from the rampant destruction of humankind. In the years since, several experiences have broadened my perspective on conservation.
I studied abroad in Botswana during college. Near the end of the semester, my class took a trip to the central Kalahari, where we saw vast landscapes filled with thriving plants and wildlife. During the drive, we stopped in a village and were introduced to a community of indigenous S¯an people. We learned that these people were being forced to leave their homelands and livelihoods, which existed within the boundaries of the Central Kalahari Game Reserve.1,2 Something struck me as fundamentally wrong about that situation. Why did the area have to support either humans or wildlife? We had just seen humans and wildlife not just surviving together but also thriving together. Removing people from the game reserve created a false dichotomy with negative ethical and existential implications for both the community we met and also humans at large, because as a species, we have historically lived in nature, rather than separated from it.
That experience started me down the path of studying plant ecology and conservation in working landscapes. I soon realized that the same issues and false dichotomies have long challenged, and are still challenging, people who live with wildlife in rural places all over the world, including in the United States. This was my first transformation—from “saving nature” to balancing conservation with livelihoods in working landscapes. Many of us working in rangeland management and rangeland science are working hard on this. We know we cannot meet our goals for conservation or for production and livelihoods by separating people and wildlife.
I started to work on livestock production and conservation issues in Kenyan savannas. At some point, I realized I had transformed into a rangeland ecologist. One thing I love about rangelands and rangeland ecology is that, by definition, they include humans and management decisions. Ecology is already complex, but when you add in the complexity of land management and human decision-making, it gets even more interesting. In Kenya, people had been managing livestock in the context of diverse wildlife assemblages for quite some time. As I studied interactions among plants, livestock management, and wildlife there, I learned there could be powerful win-wins between livestock production and conservation. More often than not, there is untapped potential for win-wins in certain places, at certain times, or at certain scales. However, I also learned that win-wins are not always possible. Sometimes, the trade-offs are real. It is generally true that you cannot achieve every objective everywhere, all the time.
In Kenya I studied temporary cattle corrals, or bomas. In eastern and southern Africa, herders have for centuries been keeping livestock in thorn-fence bomas at night to protect them from predation and theft (Figure 1 below).3, 4 Livestock graze in areas around the boma during the day. Traditional bomas range from 10 to > 100 m (32.8 to > 328 feet) in diameter and are used for months or years before being abandoned.5,6 The accumulated dung layer, which can be more than 50 centimeters (0.33 feet) deep, results in large amounts of nitrogen, phosphorus, organic carbon, and other nutrients being concentrated at abandoned sites.5, 7, 8 I want to stress that the concentration of livestock does not somehow magically create a nutrient-rich, productive site – livestock are removing nutrients from the surrounding area and concentrating them into one place. After abandonment, bomas develop into ecological hotspots characterized by high nutrient availability, high productivity, unique plant communities, and preferential use by wild life; some wild life even rely on these glades to meet critical nutrient requirements.8-12 In turn, herbivores maintain the vegetation and high nutrient status of glades through feedbacks with the plant community.7, 13-15 In central Kenya, these treeless, highly productive “glades” can persist for at least 50 years in an apparently semipermanent, self-sustaining state.5, 7 This is a win-win—livestock management creates glades that support both production and biodiversity.

Given that, we should create glades everywhere, right? Boma density is already increasing in East Africa. The impact of a boma extends beyond the physical boundaries of the corral; bomas cause changes at distances out to 100 or 150 meters (328 or 492 feet) from the corral edge.10-12, 16, 17 These edge effects include more palatable and productive vegetation, unique plant species, and increased animal use, presumably resulting from livestock use patterns (e.g., central place foraging during boma use) and subsequent wildlife foraging patterns. I expected that if two bomas were placed close together, they might generate an even larger area with the same ecosystem hotspot properties as the glade itself.18 Thus, two bomas might merge together to create a large super-glade. We did a bunch of research on this—both observational and manipulative studies where we put corrals at different distances apart. What we found was really surprising. Bomas that were very close together did in fact merge. However, bomas at intermediate distances had the opposite outcome. Areas in between these sites ended up becoming very bushy, with low nutrient quality and undesirable plant species.16, 19 This is likely because cattle were foraging and trampling between the corral sites, but they were not depositing enough nutrients from surrounding areas to convert these sites into productive glades. Thus, a few hundred feet and a little less dung turned a win-win into a tradeoff. This experience taught me to be careful with extrapolating results. Rather than assuming a given relationship will hold, we need to work hard to understand the complicated relationships between herbivores and their environments.
Rangeland Science ≠ Math
This leads me into my second transformation, which was my realization that rangelands and rangeland ecology are highly complex and context dependent. They do not operate the same way as mathematics. I like to think about grasslands as tiny forests. Just because the plants are short does not mean they are homogeneous. Grasslands have diverse canopies with multiple layers and functions (Figure 2 below). They create habitat for many different species. And these tiny forests are incredibly dynamic in space and time. Working in rangelands, we cannot assume that A + B will equal C all the time, or in every place. Many of our experiences as rangeland managers and scientists support this idea. We know that what works in one place might not work in another place, and what worked in a wet year might not work in a dry year. We know the answer is usually “it depends.”

Yet, even as we understand at some level the importance of context, rangeland science seems to be obsessed with asserting that a given management strategy or a given ecological relationship should operate the same way across vast swaths of time and space. We keep searching for and being very tempted by the idea of “one size fits all.”I have heard various assertions that if everyone would just manage or graze a certain way, all would be right with the world. But if we know there are no silver bullets, why do we keep searching for them? Here are a couple examples that have convinced me of the immense complexity and context-dependence of disturbance processes in rangelands.
One issue I have been working on is the relationship between fire and cheatgrass (Bromus tectorum L.) invasion. In the Intermountain West, invasive annual grasses increase fuel loads and fuel continuity, ultimately increasing the likelihood of fire ignition and spread in ecosystems where wildfires were historically localized or infrequent.20 After fire, cover and density of cheatgrass typically increases, creating a positive feedback cycle that has caused widespread ecosystem conversions from perennial shrublands to non-native annual grasslands across hundreds of thousands of square kilometers in the western United States.21 Impacts of invasion include reduced plant diversity, degraded wildlife habitat, lost livestock productivity, altered carbon and nitrogen cycling, and increased costs associated with fighting fires and rehabilitating invaded areas.22-25
I had done some work on the fire-invasion feedback cycle in the Great Basin, and when I moved out to the western Great Plains, I wondered if cheatgrass played the same game in this region, which has a different climate and a different history of fire and grazing. While cheatgrass has been studied intensively in the Intermountain West, much less is known about its invasion east of the Rocky Mountains.26 Several studies have reported that prescribed fires in the Great Plains can actually reduce annual brome cover,27-30 though little work had focused on the effects of wildfire, and most existing studies had lumped cheatgrass together with other brome species.
I started working in the Thunder Basin region of northeast Wyoming, which is an ecotone, or boundary zone, between sagebrush steppe and Great Plains grassland ecosystems. This region is very diverse, supporting a broad array of plant and animal species, as well as rangeland agriculture and energy development. Ecotones like Thunder Basin tend to be sensitive to environmental change, and they can respond to disturbances in surprising ways.31,32 My collaborators and I asked how fire impacted cheatgrass in this ecotone, both with an observational study and, more recently, with experimental manipulations. We found that historical wildfires had very little effect on cheatgrass—if anything, cheatgrass cover was slightly lower in burned areas than unburned areas.33 So far, our manipulative experiment also supports this result.34 Why does cheatgrass apparently behave differently in the Great Plains than in the Intermountain West? It makes sense if we think about spatial context. In the Great Plains, more summer precipitation may favor native perennials over winter annuals like cheatgrass, and native plants in the Great Plains have a longer evolutionary history of high levels of disturbance by both large ungulate herbivores and fire, so they are more tolerant of these disturbances.35,36 Thus, the relationship between cheatgrass and fire hinges on spatial context.
The story gets even more complicated if we add grazing into the mix. In my own research, I have found evidence that livestock grazing can promote invasion and it can prevent invasion. It can enhance and reduce forage production. It can enhance and reduce plant diversity. It can improve or degrade habitat quality for wildlife. Different types of grazing at the same site can produce identical outcomes or divergent outcomes. The same type of grazing applied in different places or at different times of year can generate very different outcomes.
I am not saying we know nothing. Amid all this complexity, we have managed to figure some things out. For example, we know that stocking rate is an important driver of many outcomes across many contexts.37 Rangeland managers and scientists also know a lot about how to sustainability produce livestock under wildly variable conditions.38 What I am saying is that maybe we need to focus less on what works everywhere, and more on why certain things work in certain places, or for certain objectives. We also need to embrace the idea that there are likely multiple ways to apply grazing management to achieve a given objective. In other words, we may want to spend less time searching for general rules and more time embracing and exploring the complexity within rangeland science.
For example, returning to cheatgrass, there is a lot of controversy around relationships between grazing and cheatgrass. A history of heavy grazing has been widely implicated in the spread of cheatgrass across the American West.39 We also have current examples where heavy or in some cases even moderate grazing has been linked to invasion in the Intermountain West.40-42 We have work explaining how this can happen via impacts on biocrusts or perennial plants.40,41,43,44 We also have examples from the same part of the world where moderate grazing enhanced plant community resistance to wildfire or post-fire invasion.45,46 In the western Great Plains, we have evidence from multiple studies that some level of grazing may be necessary to resist the invasion of cheatgrass and other weedy plants.47,48 In northeast Wyoming, for example, exclosures that have been keeping livestock out for over 50 years have three times as much cheatgrass as areas exposed to moderate grazing, and these long-term exclosures also have significantly fewer native plant species.49 In northern Arizona, both heavy grazing and a lack of grazing were associated with more cheatgrass than moderate grazing.50 And finally, recent work shows that it may be possible to use livestock grazing to specifically target cheatgrass. Managers may be able to graze at times of the year when cheatgrass is more palatable or more vulnerable than other desirable plants.51, 52 Alternatively, they can graze to reduce fuel loads, which can reduce wildfire risk or severity,53-56 though grazing may have minimal influence when fire weather becomes extreme.57 This new work on targeted grazing emphasizes that grazing can have dramatically different effects depending on when and how it is applied.58, 59
So what do we do with all this complexity in our science and our management? One role of rangeland science is to identify signals that cut through the noise—to look for general trends and recommendations that produce consistent outcomes and are applicable across many contexts. We cannot become paralyzed by complexity, or we will never be able to provide managers with actionable recommendations. At the same time, we cannot ignore complexity or conflicting findings; rather, I would argue that we need to be particularly attentive to the complexities inherent in our work. In cases when results do not match expectations, a common tendency is to challenge. Oh, they ignored this or that in the study. Oh, it was a dry year; it was a wet year; it was an ecotone. Oh, this result was confounded because of XYZ. All of that may be true, but I would argue there is probably a lot of useful information in every rangeland study or management trial, even and perhaps especially those conducted differently, or during a very wet or dry year. Rangeland science is still a young discipline, but we have enough information now that we can and should expand our efforts to synthesize across studies. Rather than writing off findings that do not fit our current worldview, we should challenge ourselves to broaden our views in ways that reconcile multiple findings, or multiple truths. This type of inclusive approach may allow us to unravel and understand complexities, and ultimately provide more useful and targeted management recommendations.
Humble Science
This leads me directly to my third topic, which has to do with collaborative research. I was hired by the USDA-ARS in 2013 to build a collaborative research program in northeast Wyoming. These next lessons are summarized from a talk I co-presented in 2019 at the Ecological Society of America annual meeting with my main collaborator from that region, David Pellatz, who is the executive director of the Thunder Basin Grasslands Prairie Ecosystem Association and whose family has been ranching in the Thunder Basin ecoregion for generations. We wanted to share a little bit about the process we went through to build a collaborative research program. First, like many others, we have found that there are important benefits to collaborative research approaches.60 In particular, more perspectives usually foster more creative solutions to complex problems, and these solutions also tend to have greater relevance to stakeholders and buy-in from stakeholders. The potential for real-world impact is very high.
However, there are also some substantial challenges associated with collaborative research. For example, there are trade-offs between learning and doing. Scientists are looking for a signal in the noise. This is why we are so stubborn about doing things the same way at every site, about randomizing and replicating. But managers are dealing with the noise. They are trying to work through the peculiarities of each individual situation or site to get to a good outcome on the ground. Due to this trade-off, translating real-world problems into high quality research projects can be very difficult. The stakeholders I work with have learned that research is incredibly slow, costly, and piecemeal. On the scientist side, we have learned that doing stakeholder-driven work can sometimes mean our scientific contributions are more incremental or regional. For example, it may be important to determine if a result that has been found elsewhere is also found here.
Dave and I also identified a couple of dangers to avoid when working to build a collaborative program. One big danger is the fact that different stakeholder groups may assume research results will support their management or policy agenda. Scientists need to be clear up front that results are unknown, and there are strict ethical standards surrounding data, research, and publication. It is also important to be clear up front that in rangeland systems, a scientific study is not likely to generate the final, complete answer to a real-world problem on the first try. The typical answer is “it depends.”
What has worked for us? First, being bidirectional and collaborating at all stages of research, not just at the end or the beginning. Along these lines, one thing we try to do is continually work with stakeholders to interpret research results and evaluate them against local knowledge. I am going to give you a short example here. From 2015 to 2017, we took a bunch of data on how black-tailed prairie dogs (Cynomys ludovicianus) impacted plant biomass in the Thunder Basin ecoregion. Because prairie dogs eat plants and we know they compete with cattle for forage,61-64 we expected to find less biomass inside prairie dog colonies than outside of the colonies. However, we found no significant effect of prairie dog presence on overall biomass in any of those years.65 This was pretty curious, but I tended to trust the data, particularly because the years included a dry year, an average year, and a wet year. I took these results back to the local stakeholders and they were…unconvinced. They provided some good feedback, and my collaborators and I turned around and tried to dig deeper into the data, and we also collected more data from more sites. The first thing we noticed was that the effect of prairie dogs on biomass appeared to depend on spring precipitation. Prairie dogs dramatically reduced biomass in sites or years with dry springs, and actually increased biomass in sites or years with wet springs.65 That risk of very low forage in dry years could be really important for producers, even if total biomass is not that different on average, across years. We also looked at the data by functional group and realized that species composition was markedly different bet ween colonies and non-colony locations. Colonies had much less grass biomass, and much more biomass of short-lived forbs (Figure 3 below).
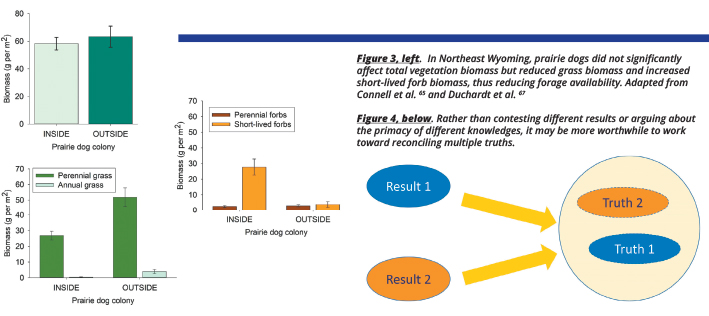
These short-lived plants may be somewhat useful to livestock early in the season, but they are a very ephemeral forage resource. The difference in composition may mean that forage on colonies is especially limiting during the dormant season, when the annuals have all disappeared and when we researchers are not usually out there measuring.
So where does this leave us? It leaves us in a place where multiple truths were reconciled through collaboration
(Figure 4 above). Yes, on average, total biomass did not differ between colonies and non-colony locations. At the same time, colonies create a big risk of forage limitation, both in dry years and during the dormant season. I probably would not have dug deeper into this issue if I had not been working closely with people in the region who pushed me to find ways of building on the initial study.
To do this type of collaborative research, the collaboration also needs to be adaptive, local, and sustained. As the real-world problems change, the focus of the research also needs to shift. Researchers need to build ties and commitment to place and understand how different stakeholders relate to place. And both scientists and stakeholders need to be ready to commit huge amounts of time and money to the collaborative process.
The prairie dog and biomass story emphasizes that on most topics, the community of ranchers, agencies, and conservation groups out there in northeast Wyoming do not actually need me to figure out how things work so they can manage better. One role of rangeland science may be to quantify and present to the outside world the things people on the ground already know. Along the way, we may come up with some surprising insights or new win-win opportunities. But I think researchers and others probably focus far too much attention on telling people how to manage and on “changing hearts and minds.” In my experience, telling people (either researchers or managers) they are doing things wrong is not typically a good strategy. What if we instead focused all that energy on listening to each other, understanding complexity, and reconciling multiple truths? It is possible we are all partly or mostly right, and we just need to figure out why and how, and in what contexts. The process of figuring this out may also allow us to move past conflict, understand the crux of the problem better, and begin moving toward actionable management solutions.
I want to come back now to the idea of humility, because this third transformation I went through was a shift in the idea of how I personally want to make change. We need scientists working across systems and at the global scale, identifying the threads that hold across space and time. But I am realizing that might not be the only way, or my favorite role. I am now more interested in digging into the complexities and communities associated with specific places. I want to really understand them and hopefully find some win-win opportunities, or at least quantify some trade-offs, for the stakeholders, livestock, and wildlife that live in or care about those places. There is a lot of value in doing research in a way that focuses on really listening to and respecting multiple perspectives so that the results we produce not only qualify as facts, but also as truths that many people can buy into and get behind. In the end, I hope this approach might help to reduce polarization in our society, especially surrounding the role of science. And maybe such an approach will also help all the world’s plants, animals, and people survive and thrive together.
Declaration of Competing Interest
I declare no competing interests.
Acknowledgments
I want to recognize the contributions of my mentors, mentees, collaborators, field technicians, landowner partners, and funders. Most of these insights came from other people, and I am very lucky to be surrounded by so many inspiring people.
Author is an Ecologist with the Rangeland Resources and Systems Research Unit, USDA Agricultural Research Service, Fort Collins, CO 80526, USA. Note: This is a written version of the plenary presentation given by the same author at the 2020 Society for Range Management meeting.
References
1. Taylor J. Celebrating San victory too soon? Reflections on the outcome of the Central Kalahari Game Reserve case. Anthropology Today. 2007; 23(5):3–5.
2. Hitchcock RK, Sapignoli M ,Babchuk WA. What about our rights? Settlements, subsistence and livelihood security among Central Kalahari San and Bakgalagadi. Int J Hum Rights. 2011; 15(1):62–88.
3. Western D, Dunne T. Environmental aspects of settlement site decisions among pastoral Maasai. Hum Ecol. 1979; 7(1):75–98.
4. Marshall F ,Reid REB, Goldstein S ,et al. Ancient herders enriched and restructured African grasslands. Nature. 2018; 561(7723):387–390.
5. Augustine DJ. Long-term, livestock-mediated redistribution of nitrogen and phosphorus in an East African savanna. J Appl Ecol. 2003; 40(1):137–149.
6. Blackmore AC ,Mentis MT ,Scholes RJ. The origin and extent of nutrient-enriched patches within a nutrient-poor savanna in South-Africa. J Biogeogr. 1990; 17(4-5):463–470.
7. Veblen KE. Savanna glade hotspots: plant community development and synergy with large herbivores. J Arid Environ. 2012; 78:119–127.
8. Reid RS, Ellis JE. Impacts of pastoralists on woodlands in south Turkana, Kenya livestock-mediated tree recruitment. Ecol Appl. 1995; 5(4):978–992.
9. Augustine DJ. Influence of cattle management on habitat selection by impala on central Kenyan rangeland. J Wildl Manag. 2004; 68(4):916–923.
10. Söderström B ,Reid RS. Abandoned pastoral settlements provide concentrations of resources for savanna birds. Acta Oecol. 2010; 36(2):184–190.
11. Vuorio V ,Muchiru A ,Reid RS, Ogutu JO. How pastoralism changes savanna vegetation: impact of old pastoral settlements on plant diversity and abundance in south-western Kenya. Biodivers Conserv. 2014; 23:3219–3240.
12. Young TP ,Patridge N ,Macrae A. Long-term glades in Acacia bushland and their edge effects in Laikipia, Kenya. Ecol Appl. 1995; 5(1):97–108.
13. Veblen KE ,Young TP. Contrasting effects of cattle and wildlife on the vegetation development of a savanna landscape mosaic. J Ecol. 2010; 98(5):993–1001.
14. Porensky LM ,Veblen KE. Grasses and browsers reinforce landscape heterogeneity by excluding trees from ecosystem hotspots. Oecologia. 2012; 168:749–759.
15. Augustine DJ, McNaughton SJ ,Frank DA. Feedbacks between soil nutrients and large herbivores in a managed savanna ecosystem. Ecol Appl. 2003; 13(5):1325–1337.
16. Porensky LM. When edges meet: interacting edge effects in an African savanna. J Ecol. 2011; 99(4):923–934.
17. Muchiru AN ,Western D ,Reid RS. The impact of abandoned pastoral settlements on plant and nutrient succession in an African savanna ecosystem. J Arid Environ. 2009; 73:322–331.
18. Porensky LM ,Young TP. Edge-effect interactions in fragmented and patchy landscapes. Conserv Biol. 2013; 27(3):509–519.
19. Porensky LM ,Young TP. Development of edge effects around experimental ecosystem hotspots is affected by hotspot density and matrix type. Landsc Ecol. 2016; 31(8):1663–1680.
20. Balch JK ,Bradley BA ,D’Antonio CM ,Gómez-Dans J. Introduced annual grass increases regional fire activity across the arid western USA (1980-2009). Global Change Biology. 2013; 19(1):173–183.
21. Bradley BA ,Curtis CA ,Fusco EJ ,et al. Cheatgrass ( Bromus tectorum) distribution in the intermountain Western United States and its relationship to fire frequency, seasonality, and ignitions. Biol Invasions. 2018; 20(6):1493–1506.
22. Rice PM. Downy Brome, Bromus tectorum L. In: Duncan CA, Clark C, eds. Invasive plants of range and wildlands and their environmental, economic, and societal impacts. Weed S cience Societ y of America; 2005:147–170.
23. DiTomaso JM. Invasive weeds in rangelands: species, impacts, and management. Weed Sci. 2000; 48(2):255–265.
24. Bradley BA, Houghton RA ,Mustard JF ,Hamburg SP. Invasive grass reduces aboveground carbon stocks in shrublands of the Western US. Glob Change Biol. 2006; 12(10):1815–1822.
25. Haferkamp MR ,Grings EE, Heitschmidt RK ,Mac-Neil MD ,Karl MG. Suppression of annual bromes impacts rangeland: animal responses. J Range Manag. 2001; 54(6):663–668.
26. Brooks ML ,Brown CS, Chambers JC ,D’Antonio CM, Keeley JE ,Belnap J. Exotic Brome-grasses in Arid and Semi-arid Ecosystems of the Western US: Causes, Consequences, and Management Implications. Exotic annual Bromus invasions: comparisons among species and ecoregions in the Western United States. S pringer; 2016.
27. Vermeire LT ,Crow der JL ,We ster DB. Plant communit y and soil environment response to summer fire in the Northern Great Plains. Rangel Ecol Manag. 2011; 64(1):37–46.
28. Whisenant SG. Postfire population dynamics of Bromus japonicus. Am Midl Nat. 1990; 123(2):301–308.
29. Harmoney KR. Grazing and burning Japanese brome ( Bromus japonicus) on mixed grass rangelands. Rangel Ecol Manag. 2007; 60(5):479–486.
30. White RS, Currie PO. Prescribed burning in the northern Great Plains: yield and cover responses of 3 forage species in the mixed grass prairie. J Range Manag. 1983; 36(2):179–183.
31. Risser PG. The status of the science examining ecotones. Bioscience. 1995; 45(5):318–325.
32. DeSantis RD, Hallgren SW ,Stahle DW. Drought and fire suppression lead to rapid forest composition change in a forest-prairie ecotone. For Ecol Manag. 2011; 261(11):1833–1840.
33. Porensky LM, Blumenthal DM. Historical wildfires do not promote cheatgrass invasion in a western Great Plains steppe. Biol Invasions. 2016; 18(11):3333–3349.
34. Estep CE . Wyoming Big Sagebrush Survival and Herbaceous Community Response to Prescribed Burns Across an Invasion Gradient of Annual Brome. MS thesis: University of Wyoming; 2020.
35. Bradley BA. Regional analysis of the impacts of climate change on cheatgrass invasion shows potential risk and opportunity.Glob Change Biol. 2009; 15(1):196–208.
36. Milchunas D ,Sala O ,Lauenroth W. A generalized model of the effects of grazing by large herbivores on grassland community structure. Am Nat. 1988; 132(1):87–106.
37. Holechek JL ,Piper RD ,Herbel CH. Range Management: Principles and Practices. 6th ed. Prentice-Hall/Pearson, Inc; 2011.
38. Espeland EK, Schreeg L ,Porensky LM. Managing risks related to climate variability in rangeland-based livestock production: what producer driven strategies are shared and prevalent across diverse dryland geographies? J Environ Manag. 2020; 255.
39. Mack RN. Invasion of Bromus tectorum L. into Western North America: an ecological chronicle. Agro-Ecosystems. 1981; 7(2):145–165.
40. Reisner MD ,Grace JB ,Pyke DA ,Doescher PS. Conditions favoring Bromus tectorum dominance of endangered sagebrush steppe ecosystems. J Appl Ecol. 2013; 50:1039–1049.
41. Holthuijzen MF ,Veblen KE. Grazing effects on precipitation-driven associations between sagebrush and perennial grasses. West N Am Nat. 2016; 76(3):313–325.
42. Williamson MA ,Fleishman E ,Mac Nally RC ,et al. Fire, livestock grazing, topography, and precipitation affect occurrence and prevalence of cheatgrass ( Bromus tectorum) in the central Great Basin, USA. Biol Invasions. 2020; 22(2):663–680.
43. Root HT, Miller JE, Rosentreter R. Grazing disturbance promotes exotic annual grasses by degrading soil biocrust communities. Ecol Appl. 2020; 30(1):e02016.
44. Slate ML ,Callaway RM ,Pearson DE. Life in interstitial space: biocrusts inhibit exotic but not native plant establishment in semi-arid grasslands. J Ecol. 2019; 107(3):1317–1327.
45. Dav ie s KW ,Bates JD ,Boyd CS ,Svejcar TJ. Prefire grazing by cattle increases postfire resistance to exotic annual grass ( Bromus tectorum) invasion and dominance for decades.Ecol Evol. 2016; 6(10):3356–3366.
46. Dav ie s KW ,Svejcar TJ ,Bates JD .Interaction of historical and nonhistorical disturbances maintains native plant communities. Ecol Appl. 2009; 19(6):1536–1545.
47. Porensky LM ,Derner JD, Augustine DJ ,Milchunas DG. Plant community composition after 75 yr of sustained grazing intensity treatments in shortgrass steppe. Rangeland Ecology & Management. 2017; 70:456–464.
48. Milchunas DG ,Lauenroth WK ,Chapman PL, Kazempour MK. Community attributes along a perturbation gradient in a shortgrass steppe. J Veg Sci. 1990; 1:375–384.
49. Porensky LM ,McGee R ,Pellatz DW. Long-term grazing removal increased invasion and reduced native plant abundance and diversity in a sagebrush grassland. Glob Ecol Conserv. 2020; 24:e01267.
50. Souther S ,Loeser M ,Crews TE ,Sisk T. Drought exacerbates negative consequences of high-intensity cattle grazing in a semiarid grassland. Ecol Appl. 2020; 30(3):e02048.
51. Schmelzer L, Perryman B ,Bruce B ,et al. Case study: reducing cheatgrass ( Bromus tectorum L.) fuel loads using fall cattle grazing. Prof Anim Sci. 2014; 30:270–278.
52. Diamond JM ,Call CA ,Devoe N. Effects of targeted grazing and prescribed burning on community and seed dynamics of a downy brome ( Bromus tectorum) dominated landscape. Invas Plant Sci Manag. 2012; 5(2):259–269.
53. Dav ie s KW ,Boyd CS ,Bates JD ,Hulet A. Winter grazing can reduce wildfire siz e,intensit y and behaviour in a shrub-grassland. Int J Wildland Fire. 2016; 25(2):191–199.
54. Dav ie s KW ,Gearhart A ,Boyd CS ,Bates JD .Fall and spring grazing influence fire ignitability and initial spread in shrub steppe communities. Int J Wildland Fire. 2017; 26(6):485–490.
55. Diamond JM ,Call CA ,Devoe N. Effects of targeted cattle grazing on fire behavior of cheatgrass-dominated rangeland in the northern Great Basin, USA. Int J Wildland Fire. 2009; 18(8):944–950.
56. Porensky LM, Perryman BL, Williamson MA ,Madsen MD, Leger EA. Combining active restoration and targeted grazing to establish native plants and reduce fuel loads in invaded ecosystems. Ecol Evol. 2018; 8(24):12533–12546.
57. Strand, Launchbaugh, Limb, Torell. Livestock grazing effects on fuel loads for wildland fire in sagebrush dominated ecosystems. J Rangel Appl. 2014; 1:35–57.
58. Bailey DW ,Mosley JC ,Estell RE ,et al. Synthesis paper: targeted livestock grazing: prescription for healthy rangelands. Rangel Ecol Manag. 2019; 72(6):865–877.
59. Dav ie s KW ,Boyd CS. Grazing is not binomial (i.e., grazed or not grazed): a reply to Herman. Bioscience. 2019; 70(1):6–7.
60. Innes JE ,Booher DE. Planning with Complexity: An Introduction to Collaborative Rationality for Public Policy. Routledge; 2010.
61. Derner JD ,Detling JK, Antolin MF. Are livestock weight gains affected by black-tailed prairie dogs? Front Ecol Environ. 2006; 4(9):459–464.
62. Koford CB. Prairie dogs, whitefaces, blue grama. Wildl Monogr. 1958; 3(3):3–78.
63. Uresk DW . Black-tailed prairie dog food habits and forage relationships in western South Dakota [Cynomys ludovicianus]. Rangel Ecol Manag/J Range Manag Arch. 1984; 37(4):325–329.
64. Miller BJ ,Reading RP, Biggins DE ,et al. Prairie dogs: an ecological review and current biopolitics. J Wildl Manag. 2007; 71(8):2801–2810.
65. Connell LC ,Porensky LM ,Scasta JD. Prairie dog (Cynomys ludovicianus) influence on forage quantity and quality in a grazed grassland-shrubland ecotone. Rangel Ecol Manag. 2019; 72(2):360–373.
66. Veblen KE ,Porensky LM. Thresholds are in the eye of the beholder: plants and wildlife respond differently to short-term cattle corrals. Bull Ecol Soc Am. 2020; 101(1):e01639.
67. Duchardt CJ ,Porensky LM, Pearse IS. Direct and indirect effects of a keystone engineer on a shrubland-prairie food web. Ecology. 2021; 102:e03195
By Lauren M. Porensky